
Zwierzęta stanowią stosunkowo małą część biomasy Ziemi, dlatego tradycyjnie nie uwzględnia się ich roli w cyklu węglowym. Modele klimatyczne całkowicie pomijają procesy z nimi związane. Jednak coraz więcej naukowców wskazuje, że może to powodować błędne oszacowania ilości węgla pobieranego i magazynowanego przez ekosystemy. Wywoływane przez zwierzęta dodatnie i ujemne sprzężenia zwrotne są bowiem w stanie, poprzez przekształcanie sieci pokarmowych, wpływać na obieg węgla w środowisku, a więc i globalne ocieplenie (Schmitz i in., 2018).
Więcej o obiegu węgla w przyrodzie – szybkim i wolnym cyklu węglowym – przeczytasz w naszych artykułach:
Dlaczego klimat się zmienia, czyli o wymuszeniach i sprzężeniach
Szybki cykl węglowy, część 1: atmosfera i ekosystemy lądowe
Szybki cykl węglowy, część 2: węgiel w oceanach
Wolny cykl węglowy i termostat węglowy

Wraz ze zmianami wielkości populacji zwierząt i wymieraniem gatunków, zaburzeniu ulegają skomplikowane zależności łączące elementy ekosystemów. Nowe szacunki tempa wymierania, włączające do wyliczeń owady i inne bezkręgowce, wskazują, że może ono obecnie sięgać 150–260 E/MSY (liczba wymarłych gatunków na 10 000 wszystkich gatunków w ciągu 100 lat), przy wartościach naturalnych kształtujących się w granicach 0,1–1 E/MSY (Cowie i in., 2022). Nie tylko pokazuje to, z jak ogromnym kryzysem mamy do czynienia, ale rodzi też pytanie, czy tak poważne zaburzenie biosfery nie wpłynie także na klimat planety. Badania wskazują, że jest to możliwe.
Zagłada megafauny
Do wymarcia plejstoceńskiej megafauny, w tym mamutów czy ogromnych leniwców, w głównej mierze przyczyniły się polowania ludzi. Część tych zwierząt było dużymi przeżuwaczami, a więc z ich obecnością wiązały się emisje metanu z fermentacji jelitowej. Naukowcy oszacowali, że wybicie wielkich przeżuwaczy spowodowało spadek globalnej temperatury Ziemi o ok. 0,08-0,2oC (Malhi i in., 2015).
1. Wieloryby, rekiny i spółka (morskie kręgowce)
Oceany pełnią niezwykle ważną rolę w cyklu węglowym. Ekosystemy morskie pobierają mniej więcej tyle samo CO2 co lądowe i usuwają z atmosfery około 1/3 antropogenicznych emisji tego gazu. Funkcjonująca w oceanach „pompa biologiczna”, której istotną częścią jest fitoplankton, pozwala wycofać część węgla z szybkiego cyklu do głębin oceanicznych i osadów, gdzie pozostaje uwięziony na setki, tysiące a nawet miliony lat (zobacz: Oceany będą pochłaniały coraz mniej dwutlenku węgla). Gdyby ten mechanizm przestał nagle działać, stężenie CO2 w atmosferze zwiększyłoby się o ok. 400 ppm w stosunku do obecnego. Dlatego wszystko, co wpływa na fitoplankton (i inne morskie organizmy fotosyntetyzujące), będzie się przekładać na możliwość działania oceanu jako „pochłaniacza” CO2. Jednym z takich czynników jest obecność i zachowanie zwierząt (Basu i Mackey, 2018, Martin i in., 2021).
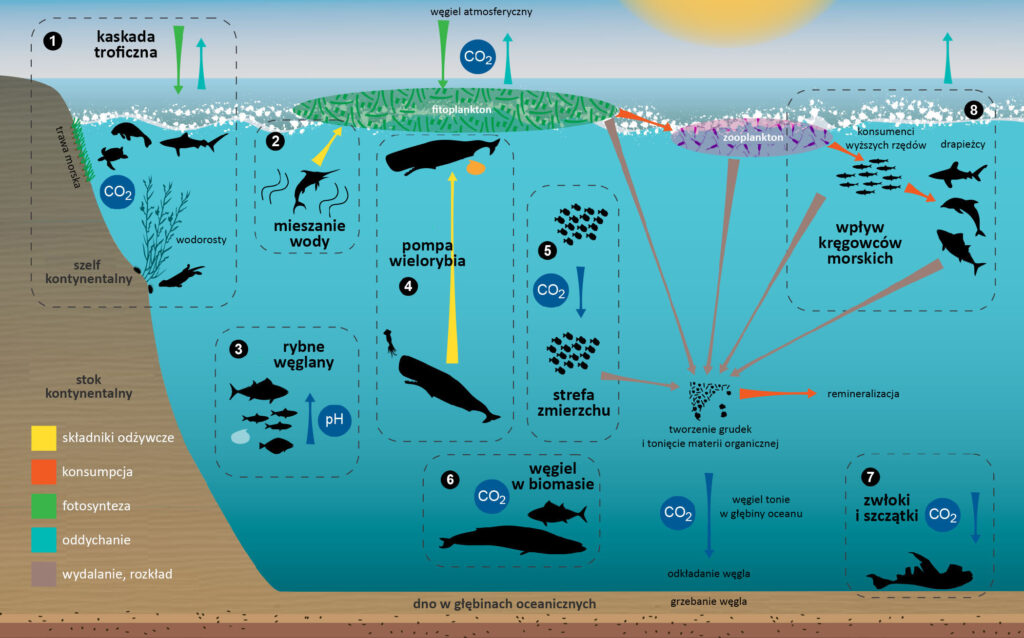
Morskie kręgowce, takie jak ryby i wieloryby, mają zarówno bezpośredni jak i pośredni wpływ na krążenie węgla. Naukowcy pod kierownictwem Angeli Martin zidentyfikowali kilkanaście takich oddziaływań i podzielili je na 4 grupy:
- związane z fizjologią (np. gromadzenie węgla w biomasie kręgowców, wydalanie z produktami ich metabolizmu),
- związane z zachowaniem (np. ruch i inne zachowania zwierząt powodują przemieszczanie się węgla wewnątrz ekosystemów i pomiędzy nimi),
- związane z odżywianiem (np. zmiany w siedliskach wynikające ze zjadania roślin, drapieżnictwo),
- „pompami składników odżywczych” (procesami powodującymi dostarczanie organizmom składników odżywczych np. azotu, umożliwiających im wiązanie węgla lub jego magazynowanie przez dłuższy czas) (Martin i in., 2021).
Żywe magazyny węgla
Węgiel wbudowany w ciała morskich kręgowców może być w nich magazynowany przez dekady lub stulecia (np. niektóre gatunki wielorybów), czyli przez podobny okres jak w lasach gospodarczych. Jeśli populacje tych zwierząt są względnie stałe, to węgiel zgromadzony w ich biomasie można uznać za zasekwestrowany właściwie na zawsze.
Niestety wielorybnictwo i rybołówstwo przyczyniają się do zmniejszania tego magazynu, nie tylko bezpośrednio, ale także pośrednio, np. gdy w poławianych populacjach maleje średni rozmiar dorosłych ryb (a więc i ilość zawartego w nich węgla organicznego). Szacuje się, że od 1950 r. morskie rybołówstwo poprzez samo pozyskiwanie tuńczyków, makreli, żaglicowatych, włócznikowatych i rekinów z głębokości poniżej 200 m zapobiegło sekwestracji około 0,02 Gt węgla (ok. 2,5 razy tyle, ile rocznie pochłaniają polskie lasy). Wynika to m.in. z faktu, że duże ryby po śmierci szybko opadają na dno, gdzie choć część materii organicznej z ich ciał może zostać zagrzebana w osadach, a więc „odcięta” od bieżącego cyklu węglowego. Ogólny potencjał morskich kręgowców, jeśli chodzi o sekwestrację węgla, szacuje się na 28-50 mln ton węgla rocznie, co odpowiada ok. ⅓-1/2 rocznych emisji Polski (Cavan i Hill, 2021, Martin i in., 2021).
Rybołówstwo wraz z wielorybnictwem wywarło też wpływ na transport węgla i innych składników odżywczych pomiędzy różnymi siedliskami. Zwierzęta migrujące mogą przenosić węgiel w biomasie i poprzez odchody na ogromne odległości. Przykładem jest tzw. „wielki taśmociąg wielorybi”. Składniki odżywcze np. azot, są transportowane przez fiszbinowce w „odpadkach” takich jak mocz, łożyska czy zwłoki pomiędzy bogatymi zazwyczaj w składniki odżywcze żerowiskami (na wysokich szerokościach geograficznych) a raczej oligotroficznymi (ubogimi) obszarami rozrodu (na niskich szerokościach). Wpływa to pozytywnie na rozwój fitoplanktonu w tych ostatnich miejscach, a tym samym zwiększa wychwytywanie CO2.

Gdyby populacja płetwali błękitnych z Oceanu Południowego osiągnęła wielkość sprzed epoki wielorybnictwa, ten efekt pozwoliłby na sekwestrację dodatkowych 0,14 mln ton węgla rocznie. Mniej więcej tyle samo mogłoby być dodatkowo magazynowane poprzez opadające na dno oceanu zwłoki wszystkich gatunków fiszbinowców, gdyby liczebność tych ssaków osiągnęła pierwotne rozmiary. Zwierzęta morskie przemieszczają składniki odżywcze również pomiędzy różnymi głębokościami (zobacz ilustracja 2, pkt 4 i 5), co także może wspomagać wiązanie CO2 w oceanach (zobacz: Oceany będą pochłaniały coraz mniej dwutlenku węgla) (Doughty i in., 2015).
Z oceanów na lądy
Niektórzy naukowcy wskazują, że zaburzenia obiegu węgla w środowisku morskim mogą odbijać się też na tym, co dzieje się na lądach. Morska i lądowa „pompy składników odżywczych” są bowiem powiązane poprzez migrujące ryby dwuśrodowiskowe, takie jak łososie, oraz poprzez ptaki morskie.
Działa to „odwrotnie” do kierunku spływu składników odżywczych z wietrzejących skał do rzek, oceanów i ostatecznie do osadów na dnie morskim (patrz Wolny cykl węglowy i termostat węglowy). Wieloryby przemieszczają bowiem te składniki z głębin oceanicznych w kierunku powierzchni (nawożą odchodami powierzchniowy fitoplankton), ryby dwuśrodowiskowe i ptaki – z powierzchni oceanu na ląd i do wód słodkich, a duże zwierzęta lądowe z biologicznych centrów, takich jak doliny rzeczne, dalej w głąb lądów.
Część gatunków zwierząt „przenosi” także składniki odżywcze między różnymi ekosystemami na lądach np. łosie i niedźwiedzie między wodnymi a leśnymi, a hipopotamy z trawiastych do słodkowodnych. Rola zwierząt może być w wielu przypadkach znacząca, np. zawartość fosforu w glebach na wyspach, gdzie obecne są ptaki morskie, jest dużo większa niż na tych, gdzie ptaków nie ma, a jeden z gatunków amazońskich małp (wełniak brunatny, Lagothrix lagotricha) jest samodzielnie odpowiedzialny za większą część transportu tego pierwiastka niż jego dostawy z nawiewanym pyłem. Ogólnie jednak efektywność “recyklingu” składników odżywczych zmalała obecnie ze względu na wymieranie zwierząt do 10% jej wartości z okresu przed zagładą megafauny na przełomie plejstocenu i holocenu. Ponieważ pierwiastki takie jak fosfor są kluczowe dla rozwoju roślin, jego braki w glebie mogą odbijać się na ich wzroście a tym samym ilości pochłanianego przez nie CO2 (Doughty i in., 2015, Malhi i in., 2015, Martin i in., 2021).
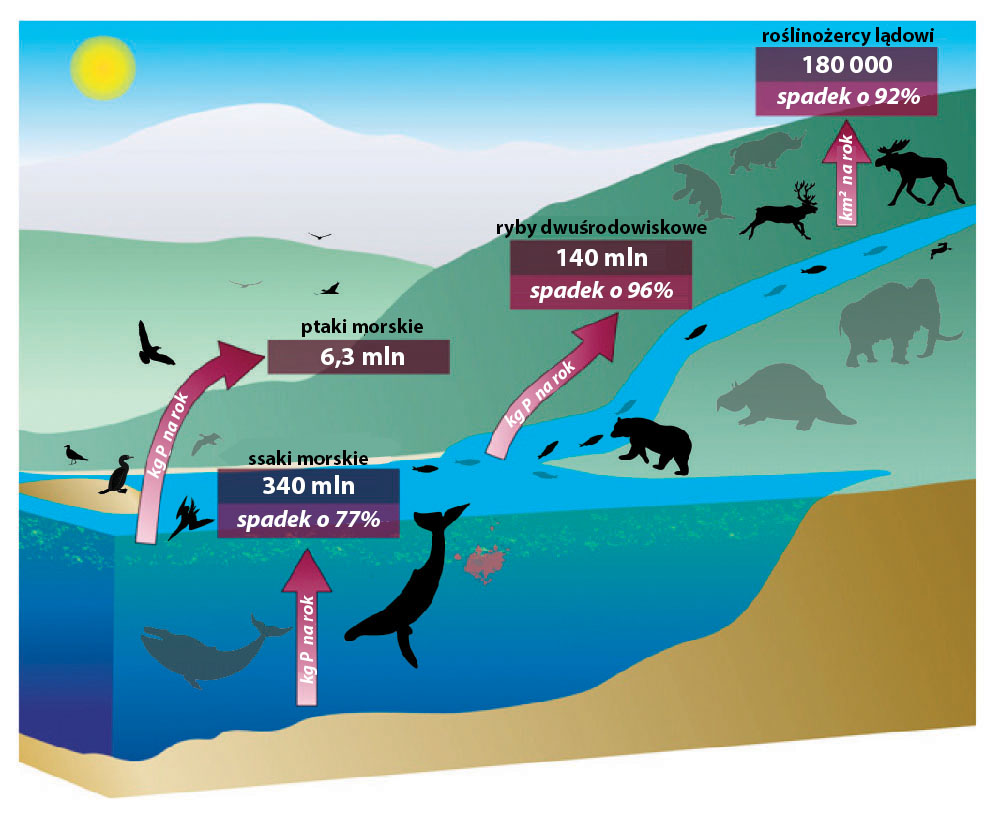
Zagrożone albatrosy i łososie
27% gatunków morskich ptaków jest sklasyfikowanych jako zagrożone, w tym 3/4 gatunków albatrosów – największych ptaków morskich. Obecna populacja ryb dwuśrodowiskowych w północno-zachodnim Pacyfiku, północno-zachodnim i północno-wschodnim Atlantyku to ok. 10% historycznej (Doughty i in., 2015).
Ryby zmniejszają zakwaszenie oceanów
Wzrost koncentracji dwutlenku węgla w atmosferze poza zmianą klimatu przynosi też drugi problem: zakwaszanie oceanów (zobacz też: 20 faktów o zakwaszaniu oceanów). Ryby doskonałokostne mogą pełnić ważną rolę w ograniczaniu skutków tego zjawiska, ich odchody są bowiem źródłem węglanu wapnia (patrz Mit: Zakwaszanie oceanu nie szkodzi morskim stworzeniom). Szacuje się, że odpowiadają za około 3-15% uwalniania tego związku w oceanach. Co więcej, z fizjologii ryb wynika, że im cieplejsza woda i więcej rozpuszczonego w niej CO2, tym więcej węglanów zawierają odchody. Niestety biorąc pod uwagę, że obecnie większość łowisk jest przełowiona lub maksymalnie wykorzystywana, ten efekt „buforowania” może być dużo niższy, niż gdyby populacje ryb były zbliżone wielkością do tych z czasów sprzed rybołówstwa przemysłowego (Martin i in., 2021).
2. Stada antylop i nosorożce (duże zwierzęta roślinożerne w tropikach)
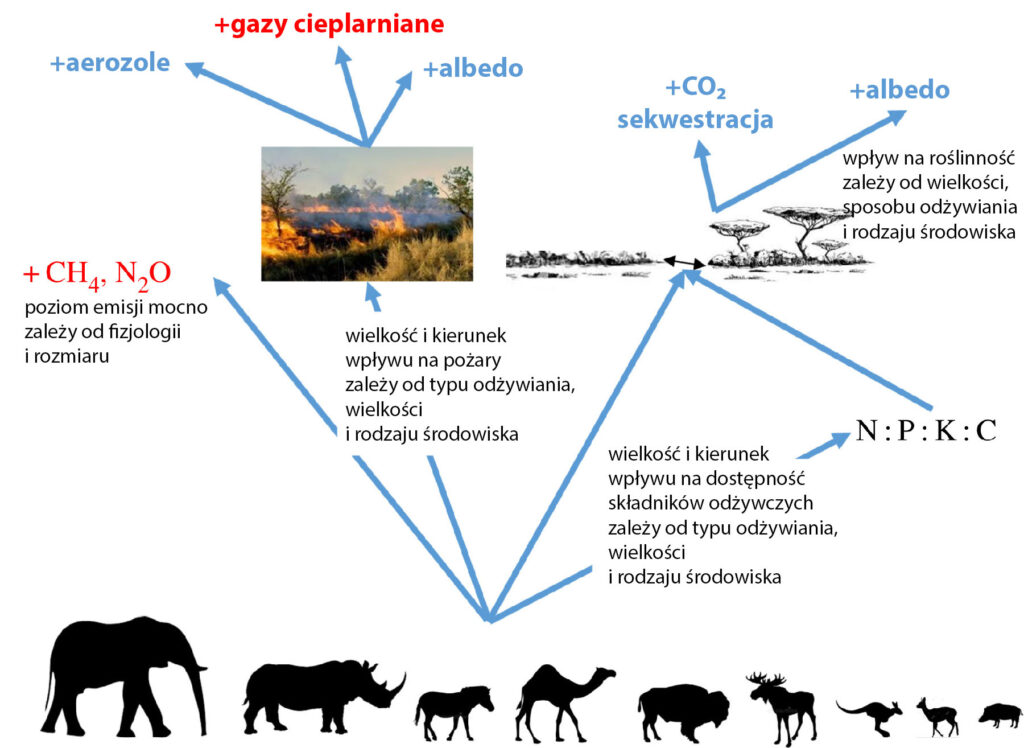
Podobnie jak morskie kręgowce, duże lądowe zwierzęta roślinożerne też biorą udział w krążeniu składników odżywczych. Trawienie liści i gałązek jest szybszą metodą przetwarzania materii organicznej niż rozkład przez mikroorganizmy glebowe, a składniki odżywcze zawarte w odchodach są łatwiej dostępne dla roślin. Dodatkowo zwierzęta transportują składniki odżywcze na spore odległości. Wpływają także na strukturę gleby wzruszając ją kopytami, rozgrzebując i ubijając, co ma znaczenie dla procesów w niej zachodzących ale może również zmieniać np. albedo (ilość odbijanego promieniowania słonecznego) danego obszaru. Od sposobu odżywiania różnych gatunków roślinożerców zależy, w dużej mierze, udział poszczególnych roślin w lokalnej florze oraz ilość gromadzących się martwych szczątków roślinnych (mogących być zarówno źródłem składników odżywczych, jak i paliwem dla pożarów).
To wszystko powoduje, że zwierzęta mogą na różne sposoby modyfikować ilość węgla gromadzonego w danym ekosystemie. Na przykład na sawannach słonie niszczą młode drzewa, co zmniejsza ilość sekwestrowanego przez nie węgla, ale jednocześnie zwiększa się albedo ze względu na „rozrzedzanie” sklepienia ciemnych drzew (Cromsigt i in., 2018, Johnson i in., 2018).
Nosorożce ograniczają pożary
Aktywność różnych gatunków zwierząt inaczej kształtuje środowisko, więc im więcej ich jest, tym bardziej zróżnicowane stają się tereny przez nie zajmowane. Badania afrykańskiej sawanny pokazały na przykład, że taka „mozaika” krajobrazowa powoduje tworzenie obszarów bardziej i mniej łatwopalnych co hamuje rozprzestrzenianie ognia.
Jakie skutki dla tych ekosystemów przynosi zmniejszanie populacji roślinożerców można było zaobserwować w latach 60. XX w. na przykładzie równiny Serengeti (Kenia/Tanzania). Populacja antylop gnu spadła wtedy gwałtownie z 1,2 mln osobników do ok. 300 000 w wyniku przeniesienia na nie choroby wirusowej bydła. To spowodowało gromadzenie się suchych, martwych traw, których było więcej, niż mogły zjeść mniejsze stada gnu i innych antylop. Dodatkowo gwałtownie wzrastająca populacja ludzi dookoła równiny zwiększała ryzyko zaprószenia ognia. W rezultacie ogromne pożary, które wtedy wybuchały, doprowadzały do wypalenia nawet 80% tego terenu, co spowodowało przekształcenie Serengeti z pochłaniacza CO2 w jego źródło. Podobna sytuacja miała miejsce w przypadku stepów azjatyckich po tym, jak praktycznie zniknęły z nich stada zwierząt takich jak suhaki, konie Przewalskiego czy gazele (Johnson i in., 2018, Schmitz i in., 2018).
Odtworzenie populacji gnu przyczyniło się do odwrócenia trendu w przypadku Serengeti. Według wyliczeń naukowców, każde dodatkowe 100 000 antylop gnu na równinie zwiększa magazynowanie węgla na tamtejszej sawannie o 15% (efekt znika po osiągnięciu pojemności środowiska). Gdy więc liczba zwierząt zbliżyła się do wartości historycznych, Serengeti ponownie stała się „pochłaniaczem” CO2 i szacuje się, że obecnie gromadzi około 1- 8 milionów ton węgla rocznie, co odpowiada od 1/5 do całości emisji ze spalania paliw kopalnych Tanzanii i Kenii.
Podobny efekt wywierają także inne gatunki zwierząt. Na sawannach szczególną rolę pełnią nosorożce, które zjadają najwyższe trawy, co ogranicza m.in. przenoszenie pożarów na drzewa. Dodatkowo powstają w ten sposób obszary pokryte niższą trawą, która jest chętniej zjadana przez antylopy. To zmniejsza tempo gromadzenia się martwych szczątków roślinnych, mogących służyć jako „paliwo” dla pożarów (Cromsigt i in., 2018, Schmitz i in., 2018).
Obserwacje prowadzone w parku Yellowstone (USA) pokazały, że na terenach zajmowanych przez jelenie (Cervus canadensis) ilość ściółki i martwej trawy stanowiła 1/6 tego, co na terenach bez jeleni. W przypadku australijskich lasów i zamieszkujących je torbaczy przekopujących ściółkę, ta redukcja wynosiła „tylko” 1/4, wystarczająco dużo jednak, by zmniejszyć wysokość płomieni o 3/4 i tempo rozprzestrzeniania pożarów o 1/3. W przypadku Afryki wzrost wielkości średniego wypalanego obszaru z powodu braku nosorożców wynikał z uruchamiania całej kaskady zależności prowadzącej do gromadzenia się większej ilości martwych szczątków roślinnych. Badania wykonane na terenie afrykańskich parków narodowych pokazały, że mógł być to 4-krotny wzrost (Imfolozi), a nawet 50-krotny (Hluhluwe). Duża różnica między parkami wynikała najprawdopodobniej z obecności innych dużych roślinożerców.
Ponieważ ograniczanie rozległości pożarów zmniejsza:
- ilość emitowanych w ich trakcie gazów cieplarnianych i sadzy,
- wywołany nimi spadek albedo terenu,
- ich negatywny wpływ na ilość CO2 pochłanianego przez rośliny,
- niszczenie gleby negatywnie wpływające na zawarte w niej składniki odżywcze,
to zwierzęta mogą mieć tu pewien wpływ na redukcję tempa zmiany klimatu. Dodatkowo ich obecność może łagodzić negatywne skutki pożarów dzięki np. transportowi składników odżywczych czy nasion roślin (Cromsigt i in., 2018, Johnson i in., 2018, Schmitz i in., 2018)

Na początku XX w. około 500 000 nosorożców wszystkich gatunków zamieszkiwało Afrykę i Azję. W latach 70. ta liczba spadła do ok. 70 000, a dziś jest ich w naturze około 27 000.
Coraz mniej nosorożców
Z 5 gatunków nosorożców krytycznie zagrożony jest czarny (około 5 500 osobników), jawajski (ok. 75) i sumatrzański (mniej niż 80). Liczebność indyjskiego wzrosła z ok. 600 osobników w latach 60. do mniej więcej 3,5 tys. obecnie. Podobnym sukcesem zakończyła się ochrona nosorożca białego (obecnie ok. 18 tys. osobników). W tym ostatnim przypadku chodzi jednak tylko o podgatunek południowy, podgatunek północny jest bowiem funkcjonalnie wymarły (żyją tylko 2 samice).
3. Tapiry, małpy i tukany (owocożercy w lasach tropikalnych)
Bioróżnorodność wpływa na zdolność lasów do wychwytywania CO2 na wiele sposobów. Większa różnorodność roślin oznacza większą produkcję biomasy i wyższą zawartość materii organicznej w glebie. Zależy ona w dużej mierze od gatunków zwierząt zamieszkujących dany obszar m.in. ze względu na to, że uczestniczą one w zapylaniu kwiatów i roznoszeniu nasion. W tym ostatnim przypadku dotyczy to ogółem prawie 90% gatunków drzew i krzewów. Podobnie jak w zbiorowiskach trawiastych, zwierzęta przemieszczają również na duże odległości składniki odżywcze, uwalniając je do otoczenia w postaci odchodów. Przetrawione resztki są przy tym łatwiej niż martwa materia roślinna dostępne dla organizmów takich jak bezkręgowce, bakterie czy grzyby (Sobral i in., 2017, Poulsen i in., 2021).
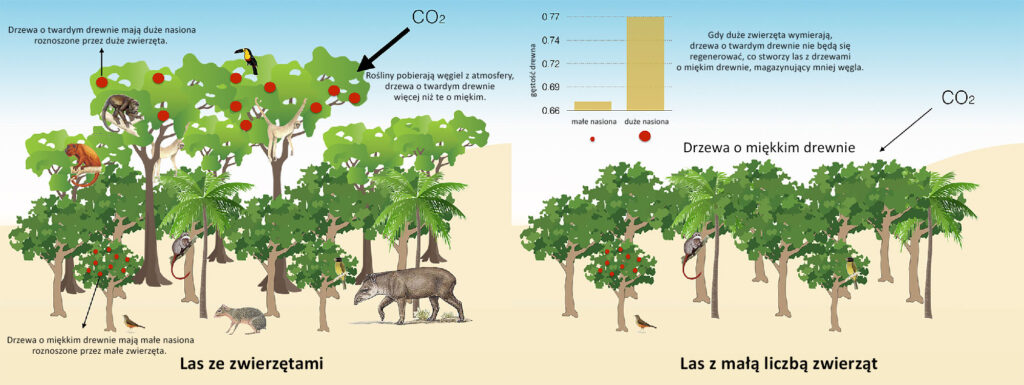
Mniej zwierząt = mniej dużych drzew
Szacuje się, że w nadziemnej biomasie lasów tropikalnych zgromadzone jest ok. 25% lądowego węgla a ich niszczenie przyczynia się do 7-17% globalnych emisji węgla. Poszczególne rosnące w nich gatunki drzew mają różne „potencjały sekwestracyjne” – im większa gęstość drewna (ciężar właściwy), średnica i wysokość dojrzałego drzewa, tym większa zdolność danego gatunku do magazynowania węgla. „Duże” gatunki mają też na ogół duże nasiona ukryte w smakowitych owocach, które są przystosowane do przenoszenia przez zwierzęta ukryte (Bello i in., 2015, Poorter i in., 2015, Sobral i in., 2017).
W Amazonii większość gatunków rozmnaża się dzięki dużym owocożercom. Jeśli w lesie jest mało zwierząt, ich sukces reprodukcyjny jest niski. Jeśli więc drzewa są wycinane lub ulegają zniszczeniu w pożarze, prawdopodobieństwo, że zastąpią je kolejne, młode drzewka jest małe. Niestety wiele owocożernych gatunków południowoamerykańskich zwierząt – takich jak tukany, czubacze, muriki (gatunek największej małpy w Ameryce Południowej), tapiry – jest zagrożonych wyginięciem. Jest to o tyle niepokojące, że gatunki drzew, które się dzięki nim rozmnażają, gromadzą około 50% węgla w Amazonii (Bello i in., 2015, Sobral i in., 2017, Schmitz i in., 2018).
Podobna sytuacja ma miejsce w lasach afrykańskich, gdzie szczególną rolę odgrywają krytycznie zagrożone słonie leśne. Zespół Fabio Berzaghiego oszacował, że ich wymarcie spowodowałoby utratę nawet ok. 7% nadziemnej biomasy lasów w centralnej Afryce, co oznacza stratę ok. 3 000 mln ton węgla. Przywrócenie zaś populacji tych słoni w dolinie Kongo do historycznego poziomu (ok. 1,1 mln osobników) pozwoliłoby zasekwestrować dodatkowo 85 mln ton węgla rocznie, co jest mniej więcej równe emisjom Francji ze spalania paliw kopalnych ( Malhi i in., 2015, Cromsigt i in., 2018, Schmitz i in., 2018, Berzaghi i in., 2019, Poulsen i in., 2021).
Duże zwierzęta – duże magazyny węgla
Duże, długowieczne zwierzęta lądowe są ponadto, podobnie jak wieloryby, same w sobie „magazynem” węgla o trwałości kilku dekad. Niestety z powodu polowań, nielegalnego handlu i utraty siedlisk co roku z tropikalnych lasów świata znikają miliony zwierząt. Gdyby całkowicie wyginęły wszystkie duże zwierzęta owocożerne (obok słoni np. też tapiry) to ekosystemy te łącznie mogłyby stracić nawet ponad 10% zgromadzonego w nich węgla (w przypadku Afryki i Ameryki Pd. łącznie, oznacza to ok. 15 mld ton węgla, czyli ok. 1,5 raza tyle co roczne antropogeniczne emisje węgla), choć na pierwszy rzut oka wydawałyby się nienaruszone. Te straty narastałyby wraz z wymieraniem zwierząt i wynikającym z tego zwiększaniem obecności w lasach gatunków roślin takich jak liany czy palmy, które słabiej gromadzą węgiel niż duże drzewa o twardym drewnie. Ochrona zwierząt owocożernych może być więc jednym ze sposobów wspomagających sekwestrację węgla w ekosystemach leśnych, w dłuższej perspektywie nawet bardziej efektywnym niż po prostu sadzenie drzew (Bello i in., 2015, Osuri i in., 2016, Cromsigt i in., 2018, Schmitz i in., 2018).
Ogromne owoce
Część drzew w Ameryce Południowej ma naprawdę ogromne owoce, zbyt duże dla współcześnie żyjących owocożerców. To gatunki, których nasiona przenosiły zwierzęta takie jak ogromne naziemne leniwce (Eremotherium). Przetrwały do obecnych czasów mimo że wymarły zwierząta, które były kluczowe dla ich rozmnażania. Naukowcy spekulują, że było to możliwe dzięki przenoszeniu owoców przez wodę, gryzonie lub ludzi, jednak zarówno zasięg występowania, zagęszczenie i zróżnicowanie genetyczne populacji tych drzew uległy dużemu zmniejszeniu (Guimarães Jr. i in., 2008, Malhi i in., 2015).
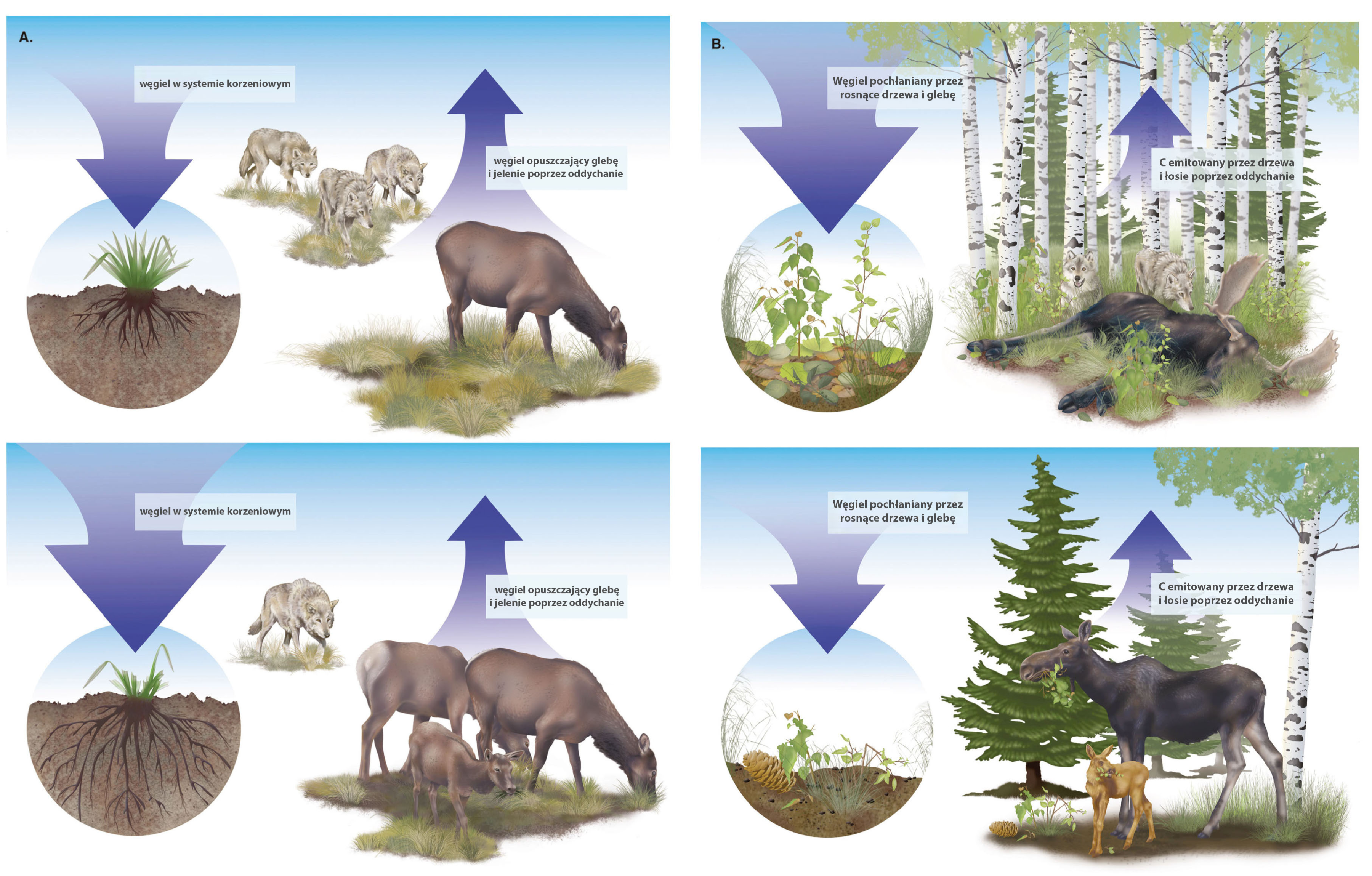
4. Renifery i łosie (duże zwierzęta roślinożerne północy)
W plejstocenie na terenie obecnej Syberii żyły ogromne stada roślinożerców takich jak mamuty, nosorożce włochate, żubry czy konie. Ich obecność, m.in. z powodu ich wpływu na krążenie składników odżywczych, była kluczowa dla powstania i utrzymania tzw. stepu mamuciego – rozległych, produktywnych ekosystemów trawiastych. W zimnym środowisku Syberii materia organiczna ulega rozkładowi w bardzo wolnym tempie, trawienie przez zwierzęta pozwala więc szybciej uzupełniać w glebie zasoby pierwiastków potrzebnych roślinom do wzrostu. Wymarcie syberyjskiej megafauny doprowadziło do przekształcenia tego regionu w środowisko uboższe, pokryte mokradłami i tajgą, z materią organiczną uwięzioną w dużej mierze w zmarzlinie.
Obecność stad zwierząt ma wpływ także m.in. na albedo obszaru Syberii, magazynowanie węgla czy temperaturę gleby. Eksperymenty prowadzone w tzw. Parku Plejstoceńskim, gdzie wprowadzono dużo roślinożerców takich jak woły piżmowe czy konie jakuckie, pokazały, że zadeptywanie śniegu przez zwierzęta może prowadzić do obniżenia temperatury gleby nawet o 15-20°C, co zmniejsza tempo rozkładu szczątków znajdujących się w zmarzlinie. Pozwala to zatrzymać w glebie materię organiczną gromadzoną przez tysiące lat, podczas których dominującymi procesami na Syberii było zatapianie szczątków na obszarach podmokłych (Doughty i in., 2015, Malhi i in., 2015, Andriuzzi i Wall, 2018, Cromsigt i in., 2018).

Liściaste czy iglaste?
W tundrze, w związku z ocieplaniem klimatu, coraz częściej obserwuje się wkraczanie krzewów i drzew. Wprawdzie rośliny drzewiaste pochłaniają więcej węgla niż niskie krzewinki tundry, jednak jednocześnie są ciemniejsze, co zmniejsza albedo tych terenów (zwiększa się ilość pochłanianego przez nie promieniowania słonecznego). W ten sposób neutralizowane są klimatyczne korzyści płynące ze zwiększonego pochłaniania CO2 przez drzewa. Zarastanie tundry jest więc przykładem dodatniego sprzężenia klimatycznego (patrz Dlaczego klimat się zmienia, czyli o wymuszeniach i sprzężeniach), nasilającego ocieplanie się klimatu. Renifery (w Ameryce Pn. zwane karibu) i woły piżmowe, które chętnie żywią się młodymi krzewami i drzewami, ograniczają ten efekt i spowalniają ocieplenie (Bakker i Svenning, 2018, Cromsigt i in., 2018)
Podobne zjawiska obserwuje się w tajdze. Na przykład łosie unikają zgryzania świerka białego, w wyniku czego tworzą się raczej rzadkie lasy świerkowe niż gęste lasy liściaste, które dominowałyby przy braku dużych roślinożerców. Wiosną, ciemniejsze tereny pokryte zwartym lasem absorbują więcej promieniowania słonecznego niż obszary, gdzie jest więcej śniegu (np. rzadki las czy otwarte przestrzenie porośnięte trawą). Obecność zwierząt także tu może mieć więc pozytywny wpływ na utrzymanie wyższego albedo i ten efekt może mieć sumarycznie większe znaczenie dla klimatu Ziemi niż ewentualne zwiększone pochłanianie CO2 przez gęstsze lasy.
Z drugiej strony badania pokazują, że obecność łosi w tajdze może spowodować zmniejszenie nawet o połowę ilości węgla magazynowego w nadziemnej biomasie roślin. Mniejsza liczba drzew liściastych oznacza także mniej szczątków opadających na dno lasu, a tym samym mniej składników odżywczych np. azotu. Latem, na terenach otwartych bardziej też nagrzewa się gleba, co może prowadzić do zwiększonej aktywności mikroorganizmów, a tym samym większych emisji CO2 i CH4. W przypadku terenów pokrytych zmarzliną ma to duże znaczenie dla regionalnego „bilansu węglowego” (Malhi i in., 2015, Wilmers i Schmitz, 2016, Bakker i Svenning, 2018, Schmitz i in., 2018).
To wszystko powoduje, że dokładne wyliczenie „klimatycznych” korzyści czy strat, które przynoszą roślinożercy na północy globu nie jest łatwe. W dużej mierze zależy ono też od wielkości populacji poszczególnych zwierząt. Dlatego istotnym elementem zapewniającym dobre funkcjonowanie ekosystemów jest obecność drapieżników.
5. Wydry, wilki i rekiny (drapieżniki)
Drapieżniki pełnią kluczową rolę w funkcjonowaniu ekosystemów zarówno lądowych jak i morskich. Ich obecność pozwala na ograniczanie populacji roślinożerców i do tego powoduje tworzenie się tzw. „krajobrazu strachu” (miejsc, których unikają roślinożercy). Dzięki temu np. lasy lepiej się odnawiają na obszarach, gdzie przebywają watahy wilków, gdyż młode drzewka są tam w mniejszym stopniu zjadane. W przypadku łosi zamieszkujących północnoamerykańską tajgę, spadek zagęszczenia ich populacji z 3 osobników na 2 km2 do 1 zwiększyłby pobieranie CO2 przez te lasy o wielkość równą mniej więcej 40% emisji Kanady ze spalania paliw kopalnych.
Jednak w ekosystemach trawiastych drapieżniki, szczególnie w zbyt dużej liczbie, mogą wywierać negatywny wpływ na gromadzenie węgla. Jak pisaliśmy w poprzednich punktach, zwierzęta takie jak jelenie stymulują wzrost traw poprzez ich zgryzanie i dodatkowo nawożą ekosystemy trawiaste odchodami, a nadmiar drapieżników może uszczuplić ich pogłowie Kuijper i in., 2013, Wilmers i Schmitz, 2016, Andriuzzi i Wall, 2018, Cromsigt i in., 2018).
Bez rekinów nie będzie podmorskich łąk
Podobne procesy obserwuje się w ekosystemach morskich. Na przykład diugonie i żółwie morskie wpływają na wzrost morskich traw, dzięki czemu wzrasta produktywność tych zbiorowisk. Odgrywają też rolę m.in. w przenoszeniu nasion części gatunków tych roślin. Jednak zbyt duża ich liczba wynikająca z braku drapieżników (np. żarłaczy tygrysich żywiących się żółwiami), może zaburzać strukturę łąk morskich traw, zmniejszać ilość szczątków opadających na dno czy naruszać osady, a przez to zmniejszać pochłanianie węgla przez te zbiorowiska.
Podobna sytuacja ma miejsce w lasach wodorostów. Wydry morskie i dorsze atlantyckie kontrolują w tych miejscach populacje jeżowców żerujących na wodorostach. Obliczenia przeprowadzone dla lasów wodorostów na zachodnim wybrzeżu Ameryki Północnej pokazały, że obecność wydr morskich może zwiększyć pobieranie CO2 w tych zbiorowiskach nawet o 1100%. Przywrócenie populacji wydr do historycznych poziomów na samym tylko wąskim pasie obszaru od wyspy Vancouver do zachodniego wybrzeża Wysp Aleuckich pozwoliłoby potencjalnie zasekwestrować 6-10% rocznych emisji Kolumbii Brytyjskiej ze spalania paliw kopalnych (Wilmers i in., 2012, Schmitz i in., 2018, Martin i in., 2021).

Glony jednokomórkowe czy wodorosty – to nie wszystko jedno
Drapieżniki mogą wywierać też jeszcze bardziej pośredni wpływ na sekwestrację. Na przykład w Bałtyku, gdy brakuje drapieżnych ryb, zakwity niektórych gatunków glonów i sinic są bardziej rozległe, co zmniejsza jakość wody i dostępność światła dla roślin morskich.
Choć i jednokomórkowe glony i trawa morska czy wodorosty to organizmy fotosyntetyzujące (czyli pochłaniają CO2), to różny jest los ich szczątków. Jednokomórkowe glony i sinice są rozkładane głównie przez bakterie blisko wybrzeży, natomiast resztki dużych roślin częściej opadają głębiej, gdzie mogą zostać zakopane w osadach. Ten proces jest odpowiednikiem sekwestracji w glebach na lądach, pozwala więc wycofywać część węgla z atmosfery na dekady czy dłużej (zobacz też: Glebowe magazyny węgla – jak je chronić przed erozją?). Gdy więc zakwity glonów zmniejszają ilość światła docierającego do wodorostów czy traw morskich, to te słabiej rosną, a tym samym mniej szczątków trafi do osadów (Schmitz i in., 2018).
Drapieżniki a słone mokradła
Podobne, skomplikowane zależności można odnotować w zbiornikach słodkowodnych, a także namorzynach, słonych bagnach czy rafach koralowych. Obserwacje prowadzone w Cape Cod (stan Massachusetts, USA) pokazały, że przełowienie drapieżnych gatunków ryb czy krabów może powodować nadmierny rozwój roślinożerców takich jak ślimaki, co prowadzi do niszczenia dużych fragmentów mokradeł i w rezultacie do erozji wybrzeża oraz wymywania odsłoniętych osadów. Powoduje to utratę węgla gromadzonego przez te ekosystemy przez setki lat a także utratę możliwości pobierania CO2 w przyszłości.
Ponieważ ekosystemy na wybrzeżach, takie jak słone mokradła, namorzyny i łąki traw morskich oraz szelfy kontynentalne, są jednymi z najważniejszych pochłaniaczy węgla na świecie, mogącymi gromadzić węgiel 40 razy szybciej od lasów tropikalnych, wszelkie procesy w nich zachodzące mogą wpływać na globalny cykl węglowy (Wilmers i in., 2012, Martin i in., 2021).

Bez dzikich zwierząt nie damy rady
To, ile węgla netto zostanie pobrane i zgromadzone w danym ekosystemie zależy od tempa fotosyntezy, strat w wyniku oddychania roślin i mikroorganizmów a także ilości materii organicznej odkładanej w glebie i osadach. Dzikie zwierzęta odgrywają ważną rolę w tych wszystkich procesach m.in. tworząc dzięki przemieszczaniu się pewną przestrzenną dynamikę. Z tego względu część naukowców proponuje, by zwiększanie populacji różnych gatunków zwierząt stało się działaniem uwzględnianym w planach mitygacji (ograniczania) zmiany klimatu (Bakker i Svenning, 2018, Cromsigt i in., 2018, Schmitz i in., 2018).
Jak wskazuje Oswald Schmitz, z Yale School for the Environment,
Odtwarzanie, wypuszczanie na wolność i ochrona funkcji jaką pełnią gatunki kręgowców i bezkręgowców może być tym, co wpłynie na sytuację [klimatyczną] poprzez zwiększenie poboru węgla o 1,5-12,5 raza (a w niektórych przypadkach więcej) w ekosystemach lądowych, słodkowodnych i morskich na świecie.
Szczególnie ważne może być przywracanie dużych gatunków roślinożernych nie będących przeżuwaczami, takich jak nosorożce. Są one obecnie krytycznie zagrożone, podobnie jak inne duże nie-przeżuwacze np. afrykański osioł nubijski czy azjatyckie baktriany, konie Przewalskiego i kulany. W pewnym stopniu mogłyby je zastąpić duże stada mniejszych, migrujących roślinożerców, jednak zjawisko takich migracji staje się coraz rzadsze (Cromsigt i in., 2018).
Zwiększanie wielkości populacji dzikich zwierząt nie jest na pewno czymś, co samodzielnie jest w stanie ograniczyć zmianę klimatu, ani głównym rozwiązaniem problemu antropogenicznych emisji gazów cieplarnianych. Może jednak być elementem strategii mitygacyjnych, szczególnie, że badania naukowe wskazują, że programy zmniejszające presję na dzikie zwierzęta czy chroniące je mogą być równie skuteczne jako działania równoważące emisje gazów cieplarnianych, jak programy sadzenia drzew typu REDD+ (Bakker i Svenning, 2018, Cromsigt i in., 2018).
Ostateczny wpływ obecności zwierząt na gromadzenie węgla jest zależny od wielu czynników i poszczególne efekty mogą wydawać się stosunkowo niewielkie. Każdy dodatkowy zasekwestrowany milion ton węgla przyczynia się jednak do opóźniania momentu przekroczenia progu 2oC globalnego ocieplenia. Biorąc pod uwagę obecną sytuację klimatyczną, nie należy odrzucać żadnych rozwiązań. Szczególnie, że odtworzenie populacji dzikich zwierząt jest metodą stosunkowo tanią i przynoszącą szereg dodatkowych korzyści np. poprawę innych niż sekwestracja węgla usług ekosystemowych.
Anna Sierpińska, konsultacja merytoryczna: dr hab. Barbara Pietrzak
Jak można zarabiać na ochronie dużych roślinożerców [ang.]: przeczytaj tutaj.
Fajnie, że tu jesteś. Mamy nadzieję, że nasz artykuł pomógł Ci poszerzyć lub ugruntować wiedzę.
Nie wiem, czy wiesz, ale naukaoklimacie.pl to projekt non-profit. Tworzymy go my, czyli ludzie, którzy chcą dzielić się wiedzą i pomagać w zrozumieniu zmian klimatu. Taki projekt to dla nas duża radość i satysfakcja. Ale też regularne koszty. Jeśli chcesz pomóc w utrzymaniu i rozwoju strony, przekaż nam darowiznę w dowolnej wysokości


